
Human Vision and Color Perception
Human stereo color vision is a very complex process that is not completely understood, despite hundreds of years of intense study and modeling. Vision involves the nearly simultaneous interaction of the two eyes and the brain through a network of neurons, receptors, and other specialized cells. The first steps in this sensory process are the stimulation of light receptors in the eyes, conversion of the light stimuli or images into signals, and transmission of electrical signals containing the vision information from each eye to the brain through the optic nerves. This information is processed in several stages, ultimately reaching the visual cortices of the cerebrum.
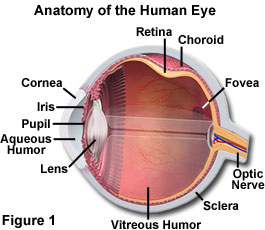
The human eye is equipped with a variety of optical components including the cornea, iris, pupil, aqueous and vitreous humors, a variable-focus lens, and the retina (as illustrated in Figure 1). Together, these elements work to form images of the objects that fall into the field of view for each eye. When an object is observed, it is first focused through the convex cornea and lens elements, forming an inverted image on the surface of the retina, a multi-layered membrane that contains millions of light-sensitive cells. In order to reach the retina, light rays focused by the cornea must successively traverse the aqueous humor (in the anterior chamber), the crystalline lens, the gelatinous vitreous body, and the vascular and neuronal layers of the retina before they reach the photosensitive outer segments of the cone and rod cells. These photosensory cells detect the image and translate it into a series of electrical signals for transmission to the brain.
Despite some misconceptions due to the wide spectrum of terminology employed for describing eye anatomy, it is the cornea, not the lens, which is responsible for the major part of the total refractive power of the eye. Being smooth and clear as glass, yet as flexible and durable as plastic, the anterior, strongly curved, transparent part of the exterior wall of the eyeball allows the image-forming light rays to pass through to the interior. The cornea also protects the eye by providing a physical barrier that shields the inside of the eye from microorganisms, dust, fibers, chemical, and other harmful materials. Although much thinner in width than the crystalline lens, the cornea provides about 65 percent of the eye's refractive power. Most of the power to bend light resides near the center of the cornea, which is rounder and thinner than the peripheral portions of the tissue.
As the window that controls the entry of light into the eye, the cornea (Figure 2) is essential to good vision and also acts as an ultraviolet light filter. The cornea removes some of the most damaging ultraviolet wavelengths present in sunlight, thereby further protecting the highly susceptible retina and crystalline lens from damage. If the cornea is curved too much, as in the case of nearsightedness, distant objects will appear as blurry images, because of imperfect light refraction to the retina. In a condition known as astigmatism, imperfections or irregularities in the cornea result in unequal refraction, which creates distortion of images projected onto the retina.
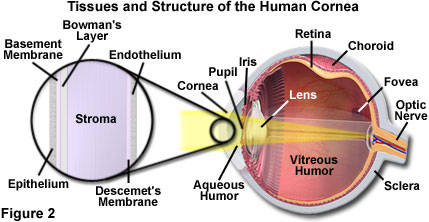
Unlike most tissues of the body, the cornea does not contain blood vessels for nourishment or to protect it against infection. Even the smallest capillaries would interfere with the precise refraction process. The cornea receives its nourishment from tears and the aqueous humor, which fills the chambers behind the structure. The outer epithelial layer of the cornea is packed with thousands of small nerve endings, making the cornea extremely sensitive to pain when rubbed or scratched. Comprising about 10 percent of the tissue's thickness, the epithelial layer of the cornea blocks foreign matter from entering the eye while providing a smooth surface for oxygen and nutrient absorption. The central layer of the cornea, known as the stroma, comprises about 90 percent of the tissue, and consists of a water-saturated fibrous protein network that provides strength, elasticity, and form to support the epithelium. Nourishing cells complete the remainder of the stroma layer. Because the stroma tends to absorb water, the endothelium tissue's primary task is to pump excess water from the stroma. Without this pumping action, the stroma would swell with water, become hazy, and ultimately turn the cornea opaque, rendering the eye blind.
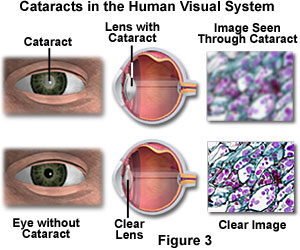
The partial or complete loss of transparency by the crystalline lens, or its capsule, results in a common condition known as cataracts. Cataracts are the leading cause of blindness worldwide and represent an important cause of visual impairment in the United States. Development of cataracts in adults is related to normal aging, sunlight exposure, smoking, poor nutrition, eye trauma, systemic disease such as diabetes and glaucoma, and undesirable side effects from some pharmaceuticals, including steroids. In the early stages, an individual suffering from cataracts perceives the world as blurry or out of focus. Clear vision is prevented by a reduction in the amount of light that reaches the retina and by clouding of the image (through diffraction and light scattering) as though the individual were observing the environment through a fog or haze (see Figure 3). Removal of the opaque lens during cataract surgery, with subsequent replacement by a plastic lens (intraocular lens implants), often results in corrected vision for unrelated conditions such as nearsightedness or farsightedness.
The function of the retina is similar to the combination of a digital image sensor (such as a charge-coupled device (CCD)) with an analog-to-digital converter, as featured in modern digital camera systems. The image-capturing receptors of the eyes, known as rods and cones, are connected with the fibers of the optic nerve bundle through a series of specialized cells that coordinate the transmission of signals to the brain. The amount of light allowed to enter each eye is controlled by the iris, a circular diaphragm that opens wide at low light levels and closes to protect the pupil (the aperture) and retina at very high levels of illumination.
As illumination changes, the diameter of the pupil (positioned in front of the crystalline lens) reflexively varies between a size of about 2 to 8 millimeters, modulating the amount of light that reaches the retina. When illumination is very bright, the pupil narrows and peripheral portions of the refractile elements are excluded from the optical pathway. The result is that fewer aberrations are encountered by image-forming light rays, and the image on the retina becomes sharper. A very narrow pupil (approximately 2 millimeters) produces diffraction artifacts that spread the image of a point source on the retina.
In the brain, the neural fibers of the optic nerves from each eye cross at the optic chiasma where visual information from both retinas traveling in parallel pathways is correlated, somewhat like the function of a time base correction generator in a digital video tape recorder. From there, the visual information travels through the optic tract to the knee-shaped lateral geniculate nuclei in the thalamus, where the signals are distributed via the optic radiations to the two visual cortices located on the lower rear section of each half of the cerebrum. In the lower layers of the cortex, the information from each eye is maintained as columnar ocular dominance stripes. As the visual signals are transmitted to the upper layers of the cortex, information from the two eyes is merged and binocular vision is formed. In abnormal ophthalmic conditions such as phorias (misalignments) of the eyes, including strabismus (better known as crossed-eyes), stereovision is disrupted as are the individual's bearings and depth perception. In cases where ophthalmic surgery is not warranted, prismatic lenses mounted in spectacles can correct some of these anomalies. Causes of interruption to the binocular fusion may be head or birth trauma, neuromuscular disease, or congenital defects.
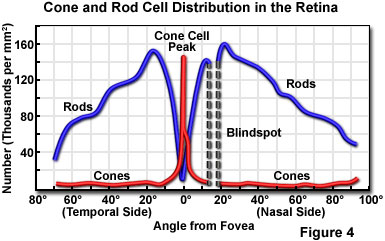
The central fovea is located in an area near the center of the retina, and positioned directly along the optical axis of each eye. Known also as the "yellow spot", the fovea is small (less than 1 square millimeter), but very specialized. These areas contain exclusively high-density, tightly packed cone cells (greater than 200,000 cones per square millimeter in adult humans; see Figure 4). The central fovea is the area of sharpest vision, and produces the maximum resolution of space (spatial resolution), contrast, and color. Each eye is populated with approximately seven million cone cells, which are very thin (3 micrometers in diameter) and elongated. The density of cone cells decreases outside of the fovea as the ratio of rod cells to cone cells gradually increases (Figure 4). At the periphery of the retina, the total number of both types of light receptors decreases substantially, causing a dramatic loss of visual sensitivity at the retinal borders. This is offset by the fact that humans constantly scan objects in the field of view (due to involuntary rapid eye movements), resulting in a perceived image that is uniformly sharp. In fact, when the image is prevented from moving relative to the retina (via an optical fixation device), the eye no longer senses an image after a few seconds.
The arrangement of sensory receptors in the outer segments of the retina partially determine the limit of resolution in different regions of the eye. In order to resolve an image, a row of less-stimulated photoreceptors must be interposed between two rows of photoreceptors that are highly stimulated. Otherwise, it is impossible to distinguish whether the stimulation originated from two closely spaced images or from a single image that spans the two receptor rows. With a center-to-center spacing ranging between 1.5 and 2 micrometers for the cones in the central fovea, optical stimuli having a separation of approximately 3 to 4 micrometers should produce a resolvable set of intensities on the retina. For reference, the radius of the first minimum for a diffraction pattern formed on the retina is about 4.6 micrometers with 550-nanometer light and a pupil diameter of 2 millimeters. Thus, the arrangement of sensory elements in the retina will determine the limiting resolution of the eye. Another factor, termed visual acuity (the ability of the eye to detect small objects and resolve their separation), varies with many parameters, including the definition of the term and the method by which acuity is measured. Over the retina, visual acuity is generally highest in the central fovea, which spans a visual field of about 1.4 degrees.
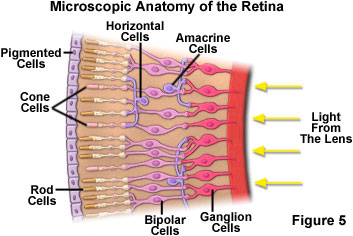
The spatial arrangement of rod and cone cells and their connection to neurons within the retina is presented in Figure 5. Rod cells, containing only the photopigment rhodopsin, have a peak sensitivity to blue-green light (wavelength of about 500 nanometers), although they display a broad range of response throughout the visible spectrum. They are the most common visual receptor cells, with each eye containing about 125-130 million rod cells. The light sensitivity of rod cells is about 1,000 times that of cone cells. However, the images generated by rod stimulation alone are relatively unsharp and confined to shades of gray, similar to those found in a black and white soft-focus photographic image. Rod vision is commonly referred to as scotopic or twilight vision because in low light conditions, shapes and the relative brightness of objects can be distinguished, but not their colors. This mechanism of dark adaptation enables the detection of potential prey and predators via shape and motion in a wide spectrum of vertebrates.
The human visual system response is logarithmic, not linear, resulting in the ability to perceive an incredible brightness range (interscene dynamic range) of over 10 decades. In broad daylight, humans can visualize objects in the glaring light from the sun, while at night large objects can be detected by starlight when the moon is dark. At threshold sensitivity, the human eye can detect the presence of about 100-150 photons of blue-green light (500 nanometers) entering the pupil. For the upper seven decades of brightness, photopic vision predominates, and it is the retinal cones that are primarily responsible for photoreception. In contrast, the lower four decades of brightness, termed scotopic vision, are controlled by the rod cells.
Adaptation of the eye enables vision to function under such extremes of brightness. However, during the interval of time before adaptation occurs, individuals can sense a range of brightness covering only about three decades. Several mechanisms are responsible for the ability of the eye to adapt to a high range of brightness levels. Adaptation can occur in seconds (by initial pupillary reaction) or may take several minutes (for dark adaptation), depending upon the level of brightness change. Full cone sensitivity is reached in about 5 minutes, whereas it requires about 30 minutes to adapt from moderate photopic sensitivity to the full scoptic sensitivity produced by the rod cells.
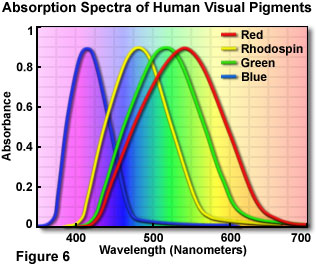
When fully light-adapted, the human eye features a wavelength response from around 400 to 700 nanometers, with a peak sensitivity at 555 nanometers (in the green region of the visible light spectrum). The dark-adapted eye responds to a lower range of wavelengths between 380 and 650 nanometers, with the peak occurring at 507 nanometers. For both photopic and scoptic vision, these wavelengths are not absolute, but vary with the intensity of light. The transmission of light through the eye becomes progressively lower at shorter wavelengths. In the blue-green region (500 nanometers), only about 50 percent of light entering the eye reaches the image point on the retina. At 400 nanometers, this value is reduced to a scant 10 percent, even in a young eye. Light scattering and absorption by elements in the crystalline lens contributes to a further loss of sensitivity in the far blue.
Cones consist of three cell types, each "tuned" to a distinct wavelength response maximum centered at either 430, 535, or 590 nanometers. The basis for the individual maxima is the utilization of three different photopigments, each with a characteristic visible light absorption spectrum. The photopigments alter their conformation when a photon is detected, enabling them to react with transducin to initiate a cascade of visual events. Transducin is a protein that resides in the retina and is able to effectively convert light energy into an electrical signal. The population of cone cells is much smaller than rod cells, with each eye containing between 5 and 7 million of these color receptors. True color vision is induced by the stimulation of cone cells. The relative intensity and wavelength distribution of light impacting on each of the three cone receptor types determines the color that is imaged (as a mosaic), in a manner comparable to an additive RGB video monitor or CCD color camera.
A beam of light that contains mostly short-wavelength blue radiation stimulates the cone cells that respond to 430-nanometer light to a far greater extent than the other two cone types. This beam will activate the blue color pigment in specific cones, and that light is perceived as blue. Light with a majority of wavelengths centered around 550 nanometers is seen as green, and a beam containing mostly 600 nanometer wavelengths or longer is visualized as red. As mentioned above, pure cone vision is referred to as photopic vision and is dominant at normal light levels, both indoors and out. Most mammals are dichromats, usually able to only distinguish between bluish and greenish color components. In contrast, some primates (most notably humans) exhibit trichromatic color vision, with significant response to red, green and blue light stimuli.
Illustrated in Figure 6 are the absorption spectra of the four human visual pigments, which display maxima in the expected red, green, and blue regions of the visible light spectrum. When all three types of cone cell are stimulated equally, the light is perceived as being achromatic or white. For example, noon sunlight appears as white light to humans, because it contains approximately equal amounts of red, green, and blue light. An excellent demonstration of the color spectrum from sunlight is the interception of the light by a glass prism, which refracts (or bends) different wavelengths to varying degrees, spreading out the light into its component colors. Human color perception is dependent upon the interaction of all receptor cells with light, and this combination results in nearly trichromic stimulation. There are shifts in color sensitivity with variations in light levels, so that blue colors look relatively brighter in dim light and red colors look brighter in bright light. This effect can be observed by pointing a flashlight onto a color print, which will result in the reds suddenly appearing much brighter and more saturated.
In recent years, consideration of human color visual sensitivity has led to changes in the long-standing practice of painting emergency vehicles, such as fire trucks and ambulances, entirely red. Although the color is intended for the vehicles to be easily seen and responded to, the wavelength distribution is not highly visible at low light levels and appears nearly black at night. The human eye is much more sensitive to yellow-green or similar hues, particularly at night, and now most new emergency vehicles are at least partially painted a vivid yellowish green or white, often retaining some red highlights in the interest of tradition.
When only one or two types of cone cells are stimulated, the range of perceived colors is limited. For example, if a narrow band of green light (540 to 550 nanometers) is used to stimulate all of the cone cells, only the ones containing green photoreceptors will respond to produce a sensation of seeing the color green. Human visual perception of primary subtractive colors, such as yellow, can arise in one of two ways. If the red and green cone cells are simultaneously stimulated with monochromatic yellow light having a wavelength of 580 nanometers, the cone cell receptors each respond almost equally because their absorption spectral overlap is approximately the same in this region of the visible light spectrum. The same color sensation can be achieved by stimulating the red and green cone cells individually with a mixture of distinct red and green wavelengths selected from regions of the receptor absorption spectra that do not have significant overlap. The result, in both cases, is simultaneous stimulation of red and green cone cells to produce a sensation of yellow color, even though the end result is achieved by two different mechanisms. The ability to perceive other colors requires the stimulation of one, two, or all three types of cone cells, to various degrees, with the appropriate wavelength palette.
Although the human visual system features three types of cones cells with their respective color pigments plus light-receptive rod cells for scotopic vision, it is the human brain that compensates for variations of light wavelengths and light sources in its perception of color. Metamers are pairs of different light spectra perceived as the same color by the human brain. Interestingly, colors that are interpreted as the same or similar by a human are sometimes readily distinguishable by other animals, most notably birds.
Intermediary neurons that ferry visual information between the retina and the brain are not simply connected one-to-one with the sensory cells. Each cone and rod cell in the fovea sends signals to at least three bipolar cells, whereas in the more peripheral regions of the retina, signals from large numbers of rod cells converge to a single ganglion cell. Spatial resolution in the outer portions of the retina is compromised by having a large number of rod cells feeding a single channel, but having many sensory cells participate in capturing weak signals significantly improves the threshold sensitivity of the eye. This feature of the human eye is somewhat analogous to the consequence of binning in slow-scan CCD digital camera systems.
The sensory, bipolar cells, and ganglion cells of the retina are also interconnected to other neurons, providing a complex network of inhibitory and excitatory pathways. As a result, the signals from the 5 to 7 million cones and 125 million rods in the human retina are processed and transported to the visual cortex by only about 1 million myelinated optical nerve fibers. The eye muscles are stimulated and controlled by ganglion cells in the lateral geniculate body, which acts as a feedback control between the retina and the visual cortex.
The complex network of excitatory and inhibitory pathways in the retina are arranged in three layers of neuronal cells that arise from a specific region of the brain during embryonic development. These circuits and feedback loops result in a combination of effects that produce edge sharpening, contrast enhancement, spatial summation, noise averaging, and other forms of signal processing, perhaps including some that have not yet been discovered. In human vision, a significant degree of image processing takes place in the brain, but the retina itself also is involved in a wide range of processing tasks.
In another aspect of human vision known as color invariance, the color or gray value of an object does not appear to change over a wide range of luminance. In 1672, Sir Isaac Newton demonstrated color invariance in human visual sensation and provided clues for the classical theory of color perception and the nervous system. Edwin H. Land, founder of the Polaroid Corporation, proposed the Retinex theory of color vision, based on his observations of color invariance. As long as color (or a gray value) is viewed under adequate lighting, a color patch does not change its color even when the luminance of the scene is changed. In this case, a gradient of illumination across the scene does not alter the perceived color or gray-level tone of a patch. If the luminance level reaches the threshold for scotopic or twilight vision, the sensation of color vanishes. In Land's algorithm, the lightness values of colored areas are computed, and the energy at a particular area in the scene is compared with all the other areas in the scene for that waveband. The calculations are performed three times, one for each waveband (long wave, short wave, and middle wave), and the resulting triplet of lightness values determines a position for the area in the three-dimensional color space defined by the Retinex theory.
The term color blindness is something of a misnomer, being widely used in colloquial conversation to refer to any difficulty in distinguishing between colors. True color blindness, or the inability to see any color, is extremely rare, although as many as 8 percent of men and 0.5 percent of women are born with some form of color vision defect (see Table 1). Inherited deficiencies in color vision are usually the result of defects in the photoreceptor cells in the retina, a neuro-membrane that functions as the imaging surface at the rear of the eye. Color vision defects can also be acquired, as a result of disease, side effects of certain medications, or through normal aging processes, and these deficiencies may affect parts of the eye other than the photoreceptors.
Normal cones and pigment sensitivity enable an individual to distinguish all the different colors as well as subtle mixtures of hues. This type of normal color vision is known as trichromacy and relies upon the mutual interaction from the overlapping sensitivity ranges of all three types of photoreceptor cone. A mild color vision deficiency occurs when the pigment in one of the three cone types has a defect, and its peak sensitivity is shifted to another wavelength, producing a visual deficiency termed anomalous trichromacy, one of three broad categories of color vision defect. Dichromacy, a more severe form of color blindness, or color deficiency, occurs when one of the pigments is seriously deviant in its absorption characteristics, or the particular pigment has not been produced at all. The complete absence of color sensation, or monochromacy, is extremely rare, but individuals with total color blindness (rod monochromats) see only varying degrees of brightness, and the world appears in black, white, and shades of gray. This condition occurs only in individuals who inherit a gene for the disorder from both parents.
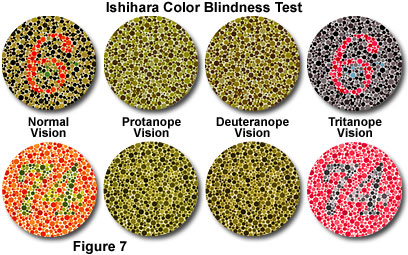
Dichromats can distinguish some colors, and are therefore less affected in their daily lives than monochromats, but they are usually aware that they have a problem with their color vision. Dichromacy is subdivided into three types: protanopia, deuteranopia, and tritanopia (see Figure 7). Approximately two percent of the male population inherits one of the first two types, with the third occurring much more rarely.
| Interactive Java Tutorial | |||||||||||
|
|||||||||||
Protanopia is a red-green defect, resulting from loss of red sensitivity, which causes a lack of perceptible difference between red, orange, yellow, and green. In addition, the brightness of red, orange, and yellow colors is dramatically reduced in comparison to normal levels. The reduced intensity effect can result in red traffic lights appearing dark (unlit), and red hues (in general), appearing as black or dark gray. Protanopes often learn to correctly distinguish between red and green, and red from yellow, primarily based on their apparent brightness, rather than on any perceptible hue difference. Green generally appears lighter than red to these individuals. Because red light occurs at one end of the visible spectrum, there is little overlap in sensitivity with the other two cone types, and people with protanopia have a pronounced loss of sensitivity to light at the long-wavelength (red) end of the spectrum. Individuals with this color vision defect can discriminate between blues and yellows, but lavender, violet, and purple cannot be distinguished from various shades of blue, due to the attenuation of the red component in these hues.
Individuals with deuteranopia, which is a loss of green sensitivity, have many of the same problems with hue discrimination as do protanopes, but have a fairly normal level of sensitivity across the visible spectrum. Because of the location of green light in the center of the visible light spectrum, and the overlapping sensitivity curves of the cone receptors, there is some response of the red and blue photoreceptors to green wavelengths. Although deuteranopia is associated with at least a brightness response to green light (and little abnormal intensity reduction), the names red, orange, yellow, and green seem to the deuteranope to be too many terms for colors that appear the same. In a similar fashion, blues, violets, purples, and lavenders are not distinguishable to individuals with this color vision defect.
Color Blindness Incidence and Causes
|
||||||||||||||||||||||||||||||||
Table 1
Tritanopia is the absence of blue sensitivity, and functionally produces a blue-yellow defect in color vision. Individuals with this deficiency cannot distinguish blues and yellows, but do register a difference between red and green. The condition is quite rare, and occurs about equally in both sexes. Tritanopes usually do not have as much difficulty in performing everyday tasks as do individuals with either of the red-green variants of dichromacy. Because blue wavelengths occur only at one end of the spectrum, and there is little overlap in sensitivity with the other two cone types, total loss of sensitivity across the spectrum can be quite severe with this condition.
When there is a loss of sensitivity by a cone receptor, but the cones are still functional, resulting color vision deficiencies are considered anomalous trichromacy, and they are categorized in a similar manner to the dichromacy types. Confusion often arises because these conditions are named similarly, but appended with a suffix derived from the term anomaly. Thus, protanomaly, and deuteranomaly produce hue recognition problems that are similar to the red-green dichromacy defects, though not as pronounced. Protanomaly is considered a "red weakness" of color vision, with red (or any color having a red component) being visualized as lighter than normal, and hues shifted toward green. A deuteranomalous individual exhibits "green weakness", and has similar difficulties in discriminating between small variations in hues falling in the red, orange, yellow, and green region of the visible spectrum. This occurs because the hues appear to be shifted toward red. In contrast, deuteranomalous individuals do not have the brightness loss defect that accompanies protanomaly. Many people with these anomalous trichromacy variants have little difficulty performing tasks that require normal color vision, and some may not even be aware that their color vision is impaired. Tritanomaly, or blue weakness, has not been reported as an inherited defect. In the few cases in which the deficiency has been identified, it is thought to have been acquired rather than inherited. Several eye diseases (such as glaucoma, which attacks the blue cones) can result in tritanomaly. Peripheral blue cone loss is most common in these diseases.
In spite of the limitations, there are some visual acuity advantages to color blindness, such as the increased ability to discriminate camouflaged objects. Outlines, rather than colors, are responsible for pattern recognition, and improvements in night vision may occur due to certain color vision deficiencies. In the military, colorblind snipers and spotters are highly valued for these reasons. During the early 1900s, in an effort to evaluate abnormal human color vision, the Nagel anomaloscope was developed. Utilizing this instrument, the observer manipulates control knobs to match two colored fields for color and brightness. Another evaluation method, the Ishihara pseudoisochromatic plate test for color blindness, named for Dr. Shinobu Ishihara, discriminates between normal color vision and red-green color blindness (as presented in the tutorial and Figure 7). A test subject with normal color vision can detect the hue difference between the figure and background. To an observer with red-green deficiency, the plates appear isochromatic with no discrimination between the figures and the design pattern.
As a natural part of the aging process, the human eye begins to perceive colors differently in later years, but does not become "colorblind" in the true sense of the term. Aging results in the yellowing and darkening of the crystalline lens and cornea, degenerative effects that are also accompanied by a shrinking of the pupil size. With yellowing, shorter wavelengths of visible light are absorbed, so blue hues appear darker. As a consequence, elderly individuals often experience difficulty discriminating between colors that differ primarily in their blue content, such as blue and gray or red and purple. At age 60, when compared to the visual efficiency of a 20-year old, only 33 percent of the light incident on the cornea reaches the photoreceptors in the retina. This value drops to around 12.5 percent by the mid-70s.
| Interactive Java Tutorial | |||||||||||
|
|||||||||||
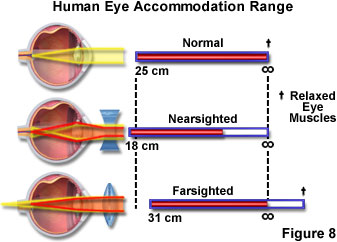
Accommodation of the eye refers to the act of physiologically adjusting the crystalline lens element to alter the refractive power and bring objects that are closer to the eye into sharp focus. Light rays initially refracted at the surface of the cornea are further converged after passing through the lens. During accommodation, contraction of the ciliary muscles relaxes tension on the lens, resulting in changes to the shape of the transparent and elastic tissue, while also moving it slightly forward. The net effect of the lens alterations is to adjust the focal length of the eye to bring the image exactly into focus onto the photosensitive layer of cells residing in the retina. Accommodation also relaxes the tension applied to the lens by the zonule fibers, and allows the anterior surface of the lens to increase its curvature. The increased degree of refraction, coupled with a slight forward shift in the position of the lens, brings objects that are closer to the eye into focus.
Focus in the eye is controlled by a combination of elements including the iris, lens, cornea, and muscle tissue, which can alter the shape of the lens so the eye can focus on both nearby and distant objects. However, in some instances these muscles do not work properly or the eye is slightly altered in shape, and the focal point does not intersect with the retina (a condition termed convergent vision). As individuals age, the lens becomes harder and cannot be properly focused, leading to poor vision. If the point of focus falls short of the retina, the condition is referred to as nearsightedness or myopia, and individuals with this affliction cannot focus on distant objects. In cases where the focal point is behind the retina, the eye will have trouble focusing on nearby objects, creating a condition known as farsightedness or hypermetropia. These malfunctions of the eye can usually be corrected with eyeglasses (Figure 8) using a concave lens to treat myopia and a convex lens to treat hypermetropia.
Convergent vision is not totally physiological and can be influenced by training, if the eyes are not defective. Repetitive procedures can be utilized to develop strong convergent vision. Athletes, such as baseball shortstops, have well-developed convergent vision. In every movement, the two eyes have to translate in unison to preserve binocular vision, with an accurate and responsive neuromuscular apparatus that is not usually subject to fatigue, controlling their motility and coordination. Changes in ocular convergence or head motion are considered in the calculations made by the complex ocular system to produce the proper neural inputs to the eye muscles. An eye movement of 10 degrees may be completed in about 40 milliseconds, with the calculations occurring faster than the eye can reach its intended target. Small eye movements are known as saccades and the larger movements from one point to another are termed versions.
The human visual system must not only detect light and color, but as an optical system, must be able to discern differences among objects, or an object and its background. Known as physiological contrast or contrast discrimination, the relationship between the apparent brightness of two objects that are seen either at the same time (simultaneous contrast) or sequentially (successive contrast) against a background, may or may not be the same. In the human visual system, contrast is reduced in environmental darkness and with individuals suffering from color visual deficiencies such as red-green color blindness. Contrast is dependent on binocular vision, visual acuity, and image processing by the visual cortex of the brain. An object with low contrast, which cannot be distinguished from the background unless it is moving, is considered camouflaged. However, colorblind individuals are often able to detect camouflaged objects because of increased rod vision and loss of misleading color cues. Increasing contrast translates into increased visibility, and a quantitative numerical value for contrast is usually expressed as a percentage or ratio. Under optimal conditions, the human eye can barely detect the presence of two percent contrast.
With human vision, an apparent increase in contrast is perceived in a narrow zone on each side of the boundary between two areas of different brightness and/or chromaticity. At the end of the nineteenth century, French physicist Michel Eug�ne Chevreul discovered simultaneous contrast. As a special function of human visual perception, the edges or contour of an object are highlighted, setting the object away from its background and easing spatial orientation. When positioned over a bright background, the region at the edge of a dark object appears lighter than the rest of the background (in effect, the contrast is enhanced). With this perception phenomenon, the color with the strongest contrast, the complementary color, is created (by the brain) at the edge. Because the color and its complement are perceived simultaneously, the effect is known as simultaneous contrast. Borders and other lines of demarcation that separate the contrasting areas tend to lessen the effect (or optical illusion) by eliminating marginal contrast. Many forms of optical microscopy, most notably phase contrast illumination, take advantage of these features of the human visual system. By increasing the physical contrast of an image without having to change the object via staining or other technique, the phase contrast specimen is protected from damage or death (in the case of living specimens).
The spatial frequency response of the human eye can be evaluated by determining the ability to detect a series of strips in a modulated sinusoidal grating. Test gratings feature alternating regions (strips) of light and dark, which increases linearly from higher to lower frequencies along the horizontal axis while contrast decreases logarithmically from top to bottom. The boundary of stripes that can just be distinguished by individuals with normal vision is between 7 and 10 cycles per degree. For achromatic vision, when the spatial frequency is very low (broad line spacing), high contrast is required to detect the sinusoidally varying intensity. As the spatial frequency rises, humans can detect periods with less contrast, reaching a peak of about 8 cycles per degree in the visual field. Beyond that point, higher contrast is again required to detect the finer sinusoidal stripes.
Examination of the modulation transfer function (MTF) of the human visual system reveals that the contrast necessary to detect the luminance variation in standardized sinusoidal gratings increases at both higher and lower spatial frequencies. In this regard, the eye behaves quite differently from a simple imaging device (such as a film camera or CCD sensor). The modulation transfer function of a simple, focused camera system displays a maximum modulation at zero spatial frequency, with the degree of modulation dropping more or less monotonically to zero at the camera's cutoff frequency.
When the luminance of a scene fluctuates periodically several times a second (as it does with television and computer monitor screens), humans perceive an irritating sensation, as though the sequential scenes were disjoined. When the fluctuation frequency increases, irritation increases and reaches a maximum at around 10 hertz, especially when bright flashes of illumination alternate with darkness. At higher frequencies, the scene no longer appears disjointed, and objects displaced from one scene to the next are now perceived to be moving smoothly. Commonly referred to as flicker, the annoying light fluttering sensation can persist up to 50-60 hertz. Beyond a certain frequency and luminance, known as the critical flicker frequency (CFF), screen flicker is no longer perceived. This is the primary reason why increasing the refresh rate of a computer monitor from 60 to 85-100 hertz produces a stable, flicker-free display.
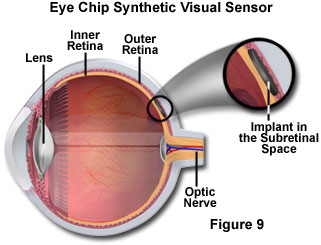
Advances in semiconductor fabrication technology, especially complementary metal oxide semiconductors (CMOS) and bipolar CMOS (BiCMOS) techniques, has led to a new generation of miniature photosensors that feature extraordinary dynamic range and fast response. Recently, arrays of CMOS sensor chips have been arranged to model the operation of the human retina. These so-called eye chips, by combining optics, human vision, and microprocessors, are advancing ophthalmology through the new field of optobionics. Damaged retinas resulting from debilitating visual diseases, such as retinitis pigmentosa and macular degeneration, as well as aging and injuries to the retina, which rob vision, are being corrected with the implanted eye chips. The silicon eye chips contain approximately 3,500 miniature light detectors attached to metal electrodes that mimic the function of the human rods and cones. The light detectors absorb incident light refracted by the cornea and lens and produce a small quantity of electrical charge that stimulates the retinal neurons. Featuring a diameter of two millimeters (see Figure 9), the replacement retina is half as thick as a typical piece of paper and is implanted into a pocket under the damaged retina.
As an alternative to the eye chip, a retinal prosthesis using a digital signal processor and a camera mounted on a pair of glasses, captures and transmits an image of an object or scene. Wirelessly, the image is sent to an embedded receiver chip near the retinal layers where nerve impulses are sent to the brain. Artificial retinas, however, will not treat glaucoma or vision deficiencies that damage the nerve fibers leading to the optic nerve. As optobionics advances, so does science's understanding of the complex human visual system.
Contributing Authors
Kenneth R. Spring - Scientific Consultant, Lusby, Maryland, 20657.
Thomas J. Fellers and Michael W. Davidson - National High Magnetic Field Laboratory, 1800 East Paul Dirac Dr., The Florida State University, Tallahassee, Florida, 32310.
BACK TO HUMAN VISION AND COLOR PERCEPTION