
Microtubules
These straight, hollow cylinders are found throughout the cytoplasm of all eukaryotic cells (prokaryotes don't have them) and carry out a variety of functions, ranging from transport to structural support. Microtubules, which are about 25 nanometers in diameter, form part of the cytoskeleton that gives structure and shape to a cell, and also serve as conveyor belts moving other organelles throughout the cytoplasm. In addition, microtubules are the major components of cilia and flagella, and participate in the formation of spindle fibers during cell division (mitosis). The length of microtubules in the cell varies between 200 nanometers and 25 micrometers, depending upon the task of a particular microtubule and the state of the cell's life cycle.
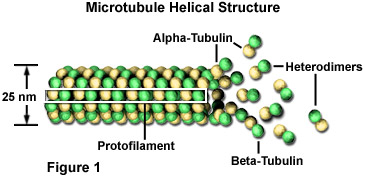
Microtubules are biopolymers that are composed of subunits made from an abundant globular cytoplasmic protein known as tubulin, as illustrated in Figure 1. Each subunit of the microtubule is made of two slightly different but closely related simpler units called alpha-tubulin and beta-tubulin that are bound very tightly together to form heterodimers. In a microtubule, the subunits are organized in such a way that they all point the same direction to form 13 parallel protofilaments. This organization gives the structure polarity, with only the alpha-tubulin proteins exposed at one end and only beta-tubulin proteins at the other.
By adding or removing globular tubulin proteins, the length of polymeric microtubules can be increased or decreased. Because the two ends of a microtubule are not the same, however, the rate at which growth or depolymerization occurs at each pole is different. The end of a polarized filament that grows and shrinks the fastest is known as the plus end and the opposing end is called the minus end. For all microtubules, the minus end is the one with exposed alpha-tubulins. In an animal cell, it is this end that is located at the centriole-containing centrosome found near the nucleus, while the plus end, comprised of exposed beta-units, is projected out toward the cell's surface. Microtubules are continuously being assembled and disassembled so that tubulin monomers can be transported elsewhere to build microtubules when needed.
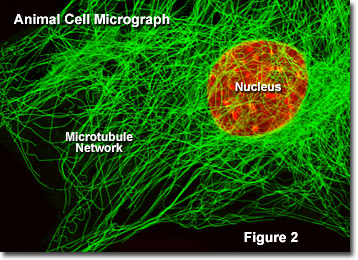
Presented in Figure 2 is a digital image of the microtubule network found in an embryonic mouse cell as seen through a fluorescence optical microscope. The extensive intertwined network is labeled with primary antibodies to alpha-tubulin, which are then stained with secondary antibodies containing a green fluorescent dye. The nucleus was counterstained with a red dye to note its location in relation to the microtubule network. Fluorescence microscopy is an important tool that scientists use to examine the structure and function of internal cellular organelles.
In addition to their structural support role, microtubules also serve as a highway system along which organelles can be transported with the aid of motor proteins. For instance, the microtubule network interconnects the Golgi apparatus with the plasma membrane to guide secretory vesicles for export, and also transports mitochondria back and forth in the cytoplasm. Another example is the translocation of vesicles containing neurotransmitters by microtubules to the tips of nerve cell axons. The motor proteins involved in organelle transport operate by altering their three-dimensional conformation using adenosine triphosphate (ATP) as fuel to move back and forth along a microtubule. With each step, the motor molecule releases one portion of the microtubule and grips a second site farther long the filament. Motor proteins, which are grouped into several distinct classes, attach to organelles through specialized receptors.
Since eukaryotic cells greatly depend upon the integrity of microtubules and other cytoskeletal filaments to maintain their structure and essentially to survive, many plants produce natural toxins aimed at disrupting the microtubule network as a means of self-defense. Taxol, for example, is a toxic substance produced by a species of yew trees that increases microtubule polymerization (building a macromolecule) by binding to the filament and stabilizing it. Other natural toxins, such as the colchicine produced by the meadow saffron, destabilize microtubules and hinder their polymerization. Both kinds of events can be fatal to the affected cell, though in some circumstances, this can be beneficial to animals, as demonstrated by taxol, which is commonly used as a cancer medication.