
Endosomes and Endocytosis
Endosomes are membrane-bound vesicles, formed via a complex family of processes collectively known as endocytosis, and found in the cytoplasm of virtually every animal cell. The basic mechanism of endocytosis is the reverse of what occurs during exocytosis or cellular secretion. It involves the invagination (folding inward) of a cell�s plasma membrane to surround macromolecules or other matter diffusing through the extracellular fluid. The encircled foreign materials are then brought into the cell, and following a pinching-off of the membrane (termed budding), are released to the cytoplasm in a sac-like vesicle. The size of vesicles varies, and those larger than 100 nanometers in diameter are typically referred to as vacuoles.
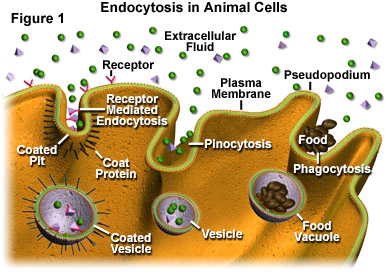
Three primary mechanisms of endocytosis that are exhibited by a typical cell are illustrated in Figure 1. On the far left of the figure, receptor mediated endocytosis, which is the most specifically-targeted form of the endocytic process, is presented. Through receptor mediated endocytosis, active cells are able to take in significant amounts of particular molecules (ligands) that bind to receptor sites extending from the cytoplasmic membrane into the extracellular fluid surrounding the cell. These receptor sites are commonly grouped together along coated pits in the membrane, which are lined on their cytoplasmic surface with a bristle-like collection of coat proteins. The coat proteins are thought to play a role in enlarging the pit and forming a vesicle. Note, as shown in Figure 1, vesicles produced via receptor mediated endocytosis may internalize other molecules in addition to ligands, though the ligands are usually brought into the cell in higher concentration.
A less specific mechanism of endocytosis is pinocytosis, which is illustrated in the central section of Figure 1. By means of pinocytosis, a cell is able to ingest droplets of liquid from the extracellular fluid. All solutes found in the droplets outside of the cell may become encased in the vesicles formed via this process, with those present in the greatest concentration in the extracellular fluid also becoming the most concentrated in the membranous sacs. Pinocytic vesicles tend to be smaller than vesicles produced by other endocytic processes.
The final type of endocytosis, termed phagocytosis (see Figure 1), is probably the most well-known manner in which a cell may import outside materials. In many school science labs, children observe amoebas under the microscope and watch the single-celled organisms eat by stretching out pseudopodia and encircling any food particles they find in their paths. This engulfment and subsequent packaging of the particles into vesicles, which are usually large enough to be correctly referred to as vacuoles, is phagocytosis. Though commonly associated with amoebas, phagocytosis is practiced by many organisms. In most multicellular animals, phagocytic cells chiefly function in bodily defense rather than as a means to gain nourishment. For example, leukocytes in the human body often phagocytose protozoa, bacteria, dead cells, and similar materials in order to help stave off infections or other problems.
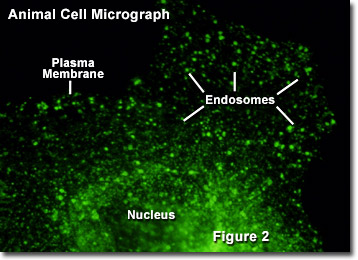
Once freed into the cytoplasm, several small vesicles produced via endocytosis may come together to form a single entity. This endosome generally functions in one of two ways. Most commonly, endosomes transport their contents in a series of steps to a lysosome, which subsequently digests the materials. In other instances, however, endosomes are used by the cell to transport various substances between different portions of the external cell membrane. This latter function is particularly important among epithelial cells, such as those that compose the outer layer of the skin, because they exhibit polarity (one side of the cell is different from the other side). Illustrated in Figure 2 is a fluorescence digital image of a single African green monkey kidney fibroblast cell (CV-1 line) transfected with a fluorescent protein fused to a targeting amino acid sequence for endosomes (green). The nucleus, plasma membrane, and endosome components are marked in the figure.
An endosome that is destined to transfer its contents to a lysosome generally goes through several changes along its way. In its initial form, when the structure is often referred to as an early endosome, the specialized vesicle contains a single compartment. Over time, however, chemical changes in the vesicle take place and the membrane surrounding the endosome folds in upon itself in a way that is similar to the invagination of the plasma membrane. In this case, however, the membrane is not pinched off. Consequently, a structure with multiple compartments, termed a multivesicular endosome, is formed. The multivesicular endosome is an intermediate structure in which further chemical changes, including a significant drop in pH level, take place as the vesicle develops into a late endosome.
Though late endosomes are capable of breaking down many proteins and fats, a lysosome is needed to fully digest all of the materials they contain. Frequently the contents of late endosomes are conveyed to a lysosome through fusion (joining together) of their membranes. Under some circumstances, late endosomes are able to further mature into lysosomes through additional chemical and structural modifications, in which case fusion is not necessary for digestion to be completed.