
Observing Mitosis with Fluorescence Microscopy
Metaphase
Perhaps the most recognizable phase of mitosis is termed metaphase, a stage where the chromosomes, attached to the kinetochore microtubules, begin to align in a single plane (known as the metaphase plate) midway between the spindle poles. The kinetochore microtubules exert tension on the chromosomes, which move back and forth in rapid erratic motion as a result, and the entire spindle-chromosome complex is now ready for the next event, separation of the daughter chromatids. Metaphase, one of the most critical stages in mitosis, occupies a substantial portion of the division cycle. The primary reason for this extended interval is that dividing cells pause until all of their chromosomes are completely aligned at the metaphase plate.
View a second, third, and fourth fluorescence image of metaphase.
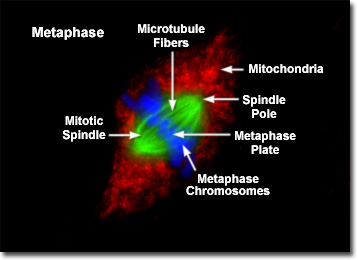
Presented in the fluorescence digital microscopy image above is a single rat kangaroo (PtK2) kidney cell in the final stages of metaphase. The chromatin is stained with a blue fluorescent probe (DAPI), while the microtubule network (mitotic spindle) is stained green (Alexa Fluor 488). Cellular mitochondria, which surround the mitotic apparatus, are stained with a red dye (MitoTracker Red CMXRos). As the dividing cell approaches metaphase, the chromosomes become highly condensed and the mitotic spindle appears (visible as a series of green tubules in the digital images of PtK2 cell metaphase). In fluorescence images of mitosis, the two poles of the spindle and the spindle fibers are clearly visible, as are the asters, which radially migrate away from the metaphase plate. The mitotic spindle contains the fibers (microtubules) responsible for translocating and separating the chromosomes.
Each spindle fiber is composed of a bundle of microtubules, which are assembled from tubulin molecules that become available as the cytoplasmic microtubular network is disassembled for cell division. The microtubules converge at each end of the spindle to a point termed the spindle pole or mitotic center. In animal cells each spindle unit contains a pair of centrioles, which are self-replicating organelles about 150 nanometers in diameter, surrounded by a radial array of aster microtubules. Plant cells, in contrast, do not contain centrioles, and their mitotic spindles are not as clearly defined at the poles. Plants lack the radial astral array characteristic of animal cells.
Fibers comprising the mitotic spindle are functionally divided into two species. The polar fibers extend to the center of the spindle pole towards the metaphase plate, while the chromosomal fibers (more commonly referred to as kinetochore fibers) travel from individual condensed chromosomes to the poles. Kinetochore fibers are attached to the chromosomes at the kinetochores, which form at specialized regions on opposite sides of the centromere. During metaphase, the chromosomes are gradually aligned at the metaphase plate, an imaginary plane located an equal distance from the poles, by forces exerted through the mitotic spindle. The chromosomes continually readjust their position to ensure that they are in the precise location for chromatid separation at the commencement of anaphase.
Protein complexes that form kinetochores in the centromere regions of sister chromatids are attached to specific repetitive DNA sequences, known as satellite DNA, which are similar in each chromosome. Mammalian centromeres form kinetochores that bind between 30 and 40 microtubules to each chromatid of the metaphase chromosome. Kinetochore microtubules from the spindle are able to traverse the nuclear region and attach to individual chromosomes when breakdown of the nuclear membrane occurs during prometaphase. Prior to this event, microtubules emanating from the spindle poles continuously probe the cytoplasm in search of kinetochores. When a kinetochore is finally captured by a microtubule, it is dragged toward the spindle pole to a region of higher microtubule density where the attachment probability is substantially increased. The kinetochore belonging to the other chromatid eventually becomes attached to microtubules from the opposite spindle pole, thus initiating final positioning of the chromosome at the metaphase plate.
Condensed chromatid pairs are held together at the metaphase plate by balanced bipolar forces exerted on the chromosomes by kinetochore microtubules in the mitotic spindle. These forces continue to operate throughout metaphase, as evidenced by the continued oscillatory motion of the chromosomes as they fine-tune their positions. Attachment of the individual chromatid kinetochores to the microtubular spindle array initiates the play of forces acting on the chromosomes. Severing one of the kinetochore microtubules from a metaphase chromosome with laser microsurgery causes the entire chromosome to immediately move to the opposite pole (to which it still remains attached). Likewise, artificially splitting the biochemical bonds between sister chromatids in metaphase will result in early migration of the chromosomes to their respective poles. Thus, the forces that act to separate chromatids in anaphase are first established when the microtubular network attaches to the kinetochores during metaphase.
BACK TO MITOSIS WITH FLUORESCENCE MICROSCOPY